USGS Groundwater Information
Why do Bacteria Colonize Aquifer Surfaces? Geochemical and Nutrient Controls of Bacterial Colonization of Silicate SurfacesBy Jennifer Roberts Rogers IntroductionIt has long been recognized that colloid-sized particles, including microorganisms, are transported through porous media (see review by Gerba and Bitton, 1984). The implications of this are of obvious importance when pathogens are involved, due to public health concerns. However, the transport of microorganisms may also have important implications of a geological or environmental nature. Microorganisms are the primary recyclers of organic carbon in aquifers, gaining energy but discarding carbon dioxide, methane, and other organic byproducts back to the groundwater. These processes can often shift geochemical equilibria by dissolving and precipitating mineral phases. In contaminated aquifers, indigenous degradative microorganisms can bioremediate groundwater contaminants, but these efforts may fail if intense bacterial attachment and growth clog sediment pores. The transport of microorganisms through porous media has long been recognized and modeled (e.g., Campbell Rehman and others, 1999). Bacterial transport in the subsurface is controlled by advection/dispersion, attachment/detachment to the solid matrix, growth phase and type of bacteria, as well as cell shape. Colloid filtration and surface interactions between bacteria and surfaces control attachment, with surface influences like electrostatic interactions, hydration forces, hydrophobic and steric interactions, and polymer bridging affecting attachment (Elimelech and others, 1995). Bacterial adhesion may be influenced by many factors including mineral and bacterial surface charges, mineral surface roughness and surface alteration, glycocalyx formation, predation, and competition. In aquifers, bacteria are often transported at a rate less than that of the flowing groundwater, leading to various representations of retardation mechanisms in models. Over the past decade, efforts have been made to characterize bacterial attachment thermodynamically through surface/solution or by using the Derjaguin-Ladau-Verwey-Overbeek (DLVO) theory to model electrostatic interactions (see review by Hermansson, 1999). Physical and hydrologic parameters also influence bacterial transport. Aquifer heterogeneities may create preferential flow paths that transport bacteria, while cell size may limit transport to larger conduits or fractures (e.g., McKay and others, 1993). Solution chemistry, including pH, DOC (dissolved organic carbon), and nutrients, is known to affect microbial attachment behavior (Mueller, 1996). Researchers have found that the attachment behavior of Pseudomonas fluorescens decreases between pH 5.5 to 7 to silica beads and with decreasing ionic strength (e.g., Deshpande and Shonnard, 1999). Field observations suggest that the number of free-living organisms increases within contaminant plumes where cells are capable of active attachment and detachment (e.g., Savage and Fletcher, 1985). In dilute or oligotrophic environments attachment may benefit microorganisms due to surface-associated organic matter. It is proposed here that microorganisms preferentially attach to surfaces that are nutritionally advantageous when those surfaces have similar electrostatic characteristics. In nutrient-limited environments, irreversible attachment of bacteria to silicate surfaces occurs predominantly to those surfaces from which the limiting nutrients (P/Fe) can be leached. In environments rich in dissolved nutrients, colonization occurs predominantly on silicate surfaces with the greatest coulobic attraction for the cells. MethodsThe study aquifer is a petroleum contaminated groundwater near Bemidji, MN, part of the USGS Toxic Substances Hydrology program. Groundwater in the study zone is completely anaerobic; a representative water has a pH of 6.6, with 3.7mmol l-1 DOC, 0.7 mmol l-1 Fe2+, and 1.0 mmol l-1 Si. The bulk solution has 4.3 mmol l-1 Ca2+, 1.5 mmol l-1 Mg2+, with 11.3 mmol l-1 HCO3- and <0.01 mmol l-1 of Al, K, Na, SO4, NO3, and PO4 (Bennett and others, 2000). Changes in groundwater chemistry in the anaerobic zone suggest silicate weathering, and evidence for accelerated dissolution of feldspars and quartz with precipitation of clay minerals has been reported (Bennett and others, 2000; 2001). Dissimilatory iron reducing bacteria (DIRB) are the dominant hydrocarbon degrading guild within the study zone, as assessed by most probable number analysis (MPN) (Bekins and others, 1999), producing high concentrations of Fe2+. Microbial colonization of silicates was characterized using the in situ microcosm method (Hiebert and Bennett, 1992). A selection of silicates were used in field experiments because they contain varying amounts of P and Fe occurring as apatite needles and iron oxides, respectively (Rogers and others, 1998) including anorthoclase (P = 1050 ppm, Fe2O3 = 4.4 %), South Dakota (S.D.) microcline (P = 1200 ppm, Fe2O3 = 0.9 %), Ontario (O.) microcline (P < 50 ppm, Fe2O3 = 1.8 %), plagioclase (P < 50 ppm, Fe2O3 = 1.0 %), and a crystalline Columbia River basalt (P = 3000 ppm, Fe2O3 = 14.0 %). The silicates were crushed and sieved to a 1-5mm size fraction, cleaned by low-power sonication and dry sterilized at 125°C. The fragments were transferred to sterile polyethylene flow-through microcosms, and suspended in the aquifer for 3 months. Additional microcosms were reserved for reference and control. Groundwater samples were taken at the time of microcosm placement and removal. Streaming potentials of mineral surfaces were measured to obtain information about grain surface charge characteristics. Samples were analyzed in a 10 mM KCl solution using a streaming potential analyzer equipped with a cylindrical cell and Ag electrodes (Brookhaven). Streaming potentials were converted to zeta potentials using the Helmholtz-Smoluchowski equation and the Fairbrother and Mastin approach (Elimelech and others, 1994). After reaction in the aquifer, microcosms were recovered anaerobically, samples were taken and immediately processed for MPN using the methods of Bekins, and others (1999). Additional samples were preserved for scanning electron microscopy (SEM). Samples were fixed in the field using chemical critical point drying (Vandevivere and Bevaye, 1993) with method blanks to insure that the surface was not altered during fixation. Surface ColonizationMicroorganisms colonize some silicate surfaces to a greater degree than others, with weathering correlating directly with the extent of colonization. Only those silicates which contain phosphorus were colonized, and this only occurred in the anaerobic groundwater (e.g., Rogers and others, 1998). SEM examination of mineral surfaces after reaction in the aquifer showed that plagioclase and the Ontario microcline (Figure 1) had little or no colonization by microorganisms and no etching. In contrast the South Dakota (SD) microcline (Figure 2) was colonized and etched.
Anorthoclase and basalt were the most heavily colonized and weathered possibly due to the presence of both P, as apatite, and Fe as iron oxides (Table 1). There is little available P in the aquifer and a strong correlation between colonization and P-content of the silicates, but the presence of ferric iron appears to intensify the degree of colonization. The anaerobic groundwater is dominated by iron reducing bacteria (DIRB) and as Fe(III) is depleted iron may become limiting as a nutrient/TEA. This suggests that microorganisms can utilize the P/Fe in silicates and further, that the increased availability of these nutrients results in microbial growth and colonization only on those surfaces. Our previous research has established that increased biomass correlates directly with the presence of P in laboratory experiments and that microorganims are capable of extracting and utilizing P from silicates (Rogers and others, 2001). This is consistent with the findings of other researchers that microorganisms derive many essential trace elements from silicate rocks (Brantley and others, 2001). While the extent of colonization correlates to the presence of P and Fe in silicates, there is no direct correlation between zeta potential and colonization. At neutral pH the feldspars and silicates used in these experiments should be uniformly negatively charged according to published pH zpc values. This is evidenced by measured zeta potentials (Table 1). The P-bearing silicates have more negative zeta potentials, yet these surfaces exhibit the greatest attachment. These results suggest that the distribution of microorganisms on different mineral surfaces in situ may be related, in part, to the nutrient content of silicates and not exclusively coulombic attraction between the cells and the mineral surfaces. However, in environments where nutrients are available or on surfaces that are positively-charged (e.g., iron oxides), it is expected that coulombic forces will dominate this interaction.
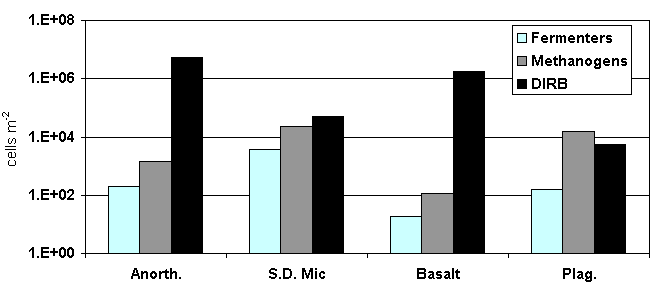
MPN analysis of the silicate chips confirms that P-bearing silicates and specifically silicates containing P and Fe support more biomass than non-nutrient silicates (Figure 3). DIRB are the most numerous physiologic type on silicate surfaces, although methanogens and fermenters are detected. Fe-bearing anorthoclase and basalt are dominated by DIRB, while SD microcline and plagioclase dislpay more balanced populations. The dominance of DIRB on Fe-bearing surfaces may indicate that these cells out-compete other metabolic guilds in the presence of iron or are actually growing and increasing their biomass once attached. Colonization on anorthoclase and basalt may be advantageous to this population by providing two necessary nutrients in one place. Because Fe(III) is used as a terminal electron acceptor (TEA) by DIRB, these organisms may have a competitive advantage when a silicate provides both P and the necessary TEA.
ImplicationsOne implication of the observed nutrient-driven colonization is that the transport of bacteria through porous media may be controlled not only by surface charge and groundwater conditions, but also by the mineral composition of the aquifer material. Small compositional changes in sediment mineralogy could have potentially tremendous impacts on which cells are transported and how fast they are transported. Conversely, the ability of bacteria to attach and grow on nutrient-rich rocks, while leaving non-nutrient rocks barren, suggests the potential for preferential clogging of the most reactive mineral surfaces and flow paths. ReferencesBekins, B.A., Godsy, E.M., and Warren, E., 1999, Distribution of microbial physiologic types in an aquifer contaminated by crude oil: Microbial Ecology, v. 37, p. 263-275. Bennett, P.C., Hiebert, F.K., and Rogers, J.R., 2000, Microbial control of mineral-groundwater equilibria - macroscale to microscale: Hydrogeology Journal, v.8, p. 47-62. Bennett, P.C., Rogers, J.R., Hiebert, F.K., and Choi, W.J., 2001, Silicates, silicate weathering, and microbial ecology.: Geomicrobiology Journal, v. 18, p. 3-19. Brantley, S.L., Lierman, L.J., Bau, M., and Wu, S., 2001, Uptake of trace metals and rare earth elements from hornblende by a soil bacterium: Geomicrobiology Journal, v. 18, p. 37-61. Campbell Rehman, L.L., Welty, C., and Harvey, R.W., 1999, Stochastic analysis of virus transport in aquifers: Water Resources Research, v. 35, no. 7, p. 1987-2006. Deshpande, P.A., and Shonnard, D.R., 1999, Modeling the effects of systematic variation in ionic strength on the attachment kinetics of Pseudomonas fluorescens UPER-1 in saturated sand columns: Water Resources Research, v. 35, p. 1619-1627. Elimelech, M., Chen, W.H., and Waypa, J.J., Desalination., Vol. 95(3), July 1994, pages 269-286., 1994, Measuring the Zeta (Electrokinetic) Potential of Reverse Osmosis Membranes by a Streaming Potential Analyzer: Desalination, v. 95, no. 3, p. 269-286. Elimelech, M., Gregory, J., Jia, X., and Williams, R., 1995, Particle Deposition and Aggregation: Measurement, Modeling and Simulation: Newton, MA, Butterworth-Heineman. Gerba, C.P., and Bitton, G., 1984, Microbial pollutants: their survival and transport pattern to groundwater, in Bitton, G., and Gerba, C.P., eds., Groundwater Pollution Microbiology: New York, John Wiley and Sons, p. 225-233. Hermansson, M., 1999, THe DLVO therory in microbial adhesion: Colloids and Surfaces, v. B14, p. 105-119. Hiebert, F.K., and Bennett, P.C., 1992, Microbial control of silicate weathering in organic-rich ground water: Science, v. 258, no. 5080, p. 278-281. McKay, L.D., Cherry, J.A., Bales, R.C., Yahya, M.T., and Gerba, C., 1993, A field example of bacteriophage as tracers in fractured flow: Environmental Science and Technology, v. 27, p. 1075-1079. Mueller, R.F., 1996, Bacterial transport and colonization in low nutrient environments: Water Research, v. 30, no. 11, p. 2681-2690. Rogers, J.R., Bennett, P.C., and Choi, W.J., 1998, Feldspars as a source of nutrients for microorganisms: American Mineralogist, v. 83, p. 1532-1540. Rogers, J.R., Bennett, P.C., and Choi, W.J., 2001, Enhanced weathering of silicates by subsurface microorganisms: a strategy to release limiting inorganic nutrients?, in Water Rock Interaction, Villasmius, Sardinia. Savage, D.C., and Fletcher, M., 1985, Bacterial Adhesion, Mechanisms, and Physiological Significance: New York, Plenum Press. Vandevivere, P., and Bevaye, P., 1993, Improved preservation of bacterial exopolymers for scanning electron microscopy: Journal of Microscopy, v. 167, p. 323-330. In George R. Aiken and Eve L. Kuniansky, editors, 2002, U.S. Geological Survey Artificial Recharge Workshop Proceedings, Sacramento, California, April 2-4, 2002: USGS Open-File Report 02-89 The use of firm, trade, and brand names in this report is for identification purposes only and does not consitute endorsement by the U.S. Government.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
![]() U.S. Department of the Interior |
U.S. Geological Survey
U.S. Department of the Interior |
U.S. Geological Survey
URL: http://water.usgs.gov/ogw/pubs/ofr0289/jrr_whydobacteria.htm
Page Contact Information: Contact the USGS Office of Groundwater
Page Last Modified: Wednesday, 28-Dec-2016 01:48:59 EST